

Бурая жировая ткань впервые была описана несколько сот лет назад. Эта ткань локализована близ жизненно важных органов и играет главную роль в производстве тепла, необходимого для поддержания постоянной температуры, в особенности у мелких животных, живущих в условиях холода и/или впадающих в зимнюю спячку, а также у новорожденных, в том числе и человека.
Нейротрансмиттер норадреналин инициирует термогенез и увеличивает термогенную активность ткани при хроническом холодовом стрессе, стимулируя её гиперплазию и гипертрофию. Норадреналин многократно ускоряет как пролиферацию клеток, так и их дифференцировку. В бурых преадипоцитах норадреналин стимулирует синтез ДНК и белка по сАМР-зависимому пути. В постконфлуентных клетках норадреналин стимулирует экспрессию гена белка-разобщителя. Была показана линейная корреляция между экспрессией гена и повышением сАМР. Таким образом, процессы; пролиферации, дифференцировки и термогенеза, опосредованы одним и тем же мессенджером – сАМР и инициируются через β-адренорецепторы. Роль α1-адренорецепторов в указанных процессах оказалась минорной и сводилась к синергическому усилению сигнала, индуцированного через β-рецепторы и сАМР. В этом случае ионы Са2+ усиливали стимулирующее действие сАМР на термогенез клеток и экспрессию ряда генов [1,2].
Единственным указанием на возможное участие ионов Са2+ в активации пролиферации бурых преадипоцитов являлась установленная ранее корреляция между эффектами нейропептидов на [Са2+]i в свежевыделенных преадипоцитах и модуляцией пролиферации культивируемых клеток пептидами при тех же концентрациях. Однако оказалось, что [Са2+]i, инициируемый норадреналином и пептидами в преадипоцитах и зрелых клетках бурого жира отличается по ряду параметров (кинетика, амплитуда и т.д.). Более того, создавалось впечатление, что Са2+-сигналы, инициируемые в преадипоцитах разными агонистами, обладают разными физиологическими функциями в этих клетках и связаны, по-видимому, со стимуляцией противоположно направленных процессов. Поэтому мы считаем весьма важным на первом этапе установить, причины существенных отличий Са2+-ответов на норадреналин в преадипоцитах и зрелых клетках бурого жира. Это позволит в последующем использовать полученные знания для контролируемого управления за развитием клеток и ткани.
К важным свойствам бурой жировой ткани относится её способность наращивать свою массу в случае хронического холодового стресса. Запуск этого процесса, также как и термогенеза, осуществляется в основном норадреналином. Бурая жировая ткань представляет собой хорошую модель для исследования механизмов гормонального контроля за развитием ткани и клеток [1-6].
Материалы и методы исследования
Эксперименты проводили на суспензии свежевыделенных бурых преадипоцитов. Мышей-самцов линии NMRI в возрасте 3-5 недель содержали в отсутствие холодового стресса (температура в помещении 20-23оС). Забор ткани производили из затылочной, межлопаточной и пазушной областей. Бурую жировую ткань помещали в стандартный солевой буфер, содержащий (в мМ): NaCl – 123, KCl – 5, CaCl – 1,3, глюкозы – 5, HEPES – 100, BSA – 1,5%, pH – 7,4. Ткань бурого жира переносили в пробирки с раствором коллагеназы для разрушения межклеточного матрикса. Раствор коллагеназы (0,7-0,8 мг/мл изоляционного буфера) готовили непосредственно перед опытом из расчета 2–2,5 мл раствора на одну мышь. Измельченную ткань инкубировали в растворе коллагеназой на водяной бане при 28оС в течение 20-30 мин, периодически встряхивая на шейкере, и затем охлаждали на льду в течение 15-30 мин. Суспензию клеток фильтровали на нейлоновом фильтре (размер пор 250 мкм) и центрифугировали течение 10 мин при 1200 g. Слой белого жира удаляли и супернатант отбирали шприцем с длинной иглой. Полученный осадок ресуспендировали в 9 мл среды DMEM с добавлением 10 мМ HEPES и BSA (0,5 мг/мл), рН 7,4, отфильтровывали на нейлоновом фильтре (размер пор 25 микрон) и центрифугировали течение 10 мин при 1200 g. Осадок ресуспендировали в среде DMEM без альбумина. Зрелые адипоциты содержат жировые капли и не осаждаются при данных параметрах центрифугирования.
Измерения [Ca2+]i проводили спектрофлуориметрически с помощью внутриклеточных ион-селективных флуоресцентных зондов Fura-2/AM. В данной работе мы регистрировали кинетики [Са2+]i при аппликации агонистов адренорецепторов в течение 20 минут и далее учитывали и анализировали величину [Са2+]i на 20-й минуте. Статистическую обработку результатов экспериментов проводили с использованием программы Origin 7,0.
Результаты исследования и их обсуждение
1. Роль α1- и β-адренорецепторов в формировании Са2+-сигнала в преадипоцитах бурого жира.
В зрелых бурых адипоцитах норадреналин (НА) и α1-селективный агонист циразолин вызывали очень схожие Ca2+-ответы, в то время как влияние изопротеренола на [Ca2+]i было на два порядка меньше, чем влияние НА и циразолина. В свежевыделенных бурых преадипоцитах эффективность указанных агонистов была совершенно иной. Агонисты α1-адренорецепторов: циразолин, фенилефрин (не показано) и оксиметазолин (агонист α1А-рецепторов, которые составляют 80-90 % от α1-адренорецепторов в бурой жировой ткани) вызывали в 2-3 раза меньшее увеличение [Ca2+]i по сравнению с НА, тогда как изопротеренол и НА инициировали в них практически идентичные Са2+-ответы (рис. 1). Агонист α2-адренорецепторов клонидин не имел эффекта в бурых преадипоцитах.
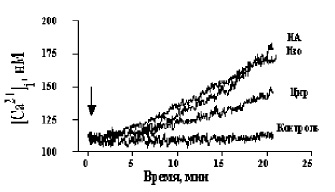
Рис. 1. Кинетика Са2+-ответов в бурых преадипоцитах, инициированных 3 mМ циразолином (Цир), 3 mМ изопротеренолом (Изо) и 3 mМ норадреналином (НА), контроль
При действии адренергических агонистов наблюдался аддитивный Са2+-ответ, стимулированный через β- и α1-адренорецепторы; [Ca2+]i при аппликации НА была примерно равна арифметической сумме эффектов изопротеренола и оксиметазолина. Совместная аппликация β- и α1-агонистов при максимальной концентрации 10 μМ последних дала в результате Са2+-ответы, аналогичные эффектам НА. Для того, чтобы подтвердить относительный вклад α1- и β-адренорецепторов в НА- иницированный Ca2+-ответ, были исследованы эффекты α1/2- и β-селективных антагонистов- фентоламина и надолола, соответственно. Надолол на 90 % подавлял НА- иницированный Ca2+-ответ, в то время как фентоламин уменьшал Са2+-ответ только на 25 % (рис.2). Таким образом, оба типа адренорецепторов (α1- и β-), вносят вклад в формирование [Ca2+]i, однако главная роль в формировании Са2+-ответа в бурых преадипоцитах принадлежит β-адренорецепторам [1-6].
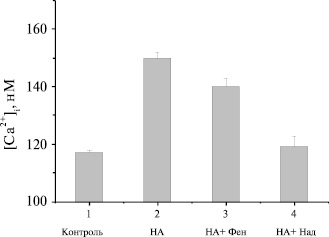
Рис. 2. Влияние селективных антагонистов α1- и β-адренорецепторов на НА-иницированный Са2+-ответ в бурых преадипоцитах: 1 – контроль; 2 – 6 mМ норадреналина (НА); 3 – НА + 10 mМ фентоламина; 4 – НА + 10 mМ надолола. (n=5). Уровень [Ca2+]I измеряли через 20 мин после добавления антагонистов
2. Действие ингибиторов и активаторов сАМР-зависимого пути на Са2+-сигнал в преадипоцитах.
Активация β-адренорецепторов клеток бурого жира приводит к опосредованной Gs-белками стимуляции аденилатциклазы (АЦ) и к образованию вторичного мессенджера сАМР. Увеличение внутриклеточного уровня сАМР можно достичь либо добавлением к клеткам проникающего через плазматическую мембрану аналога сАМР-BrcAMP, либо добавлением форсколина- непосредственного активатора АЦ. На рис. 3 представлены кинетики Са2+-ответов бурых преадипоцитов на норадреналин, BrcAMP и форсколин. Как видно из рисунка, увеличение [Са2+]i в ответ на форсколин и BrcAMP (кривые 3 и 4) заметно выше, чем при аппликации НА, что еще раз подчеркивает ведущую роль β-адренорецепторов в формировании [Са2+]i.
Добавление 500 μМ BrcAMP вызывает максимальное увеличение [Са2+]i, что следует из колоколообразной кривой дозозависимости (рис. 4).
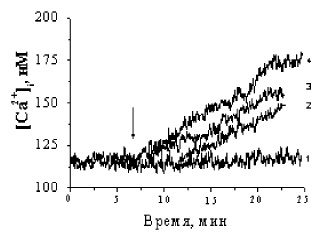
Рис 3. Изменение Са2+-ответа в свежевыделенных преадипоцитах под влиянием норадреналина, BrcAMP и форсколина: 1 – контроль; 2 – 1 μМ норадреналин; 3 – 10 μМ форсколин; 4 – 0,5 μМ BrcAMP
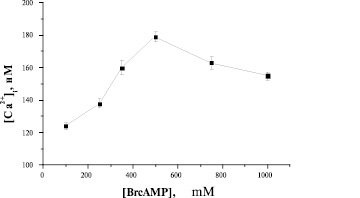
Рис. 4. Зависимость Са2+-ответа свежевыделенных преадипоцитов от концентрации BrcAMP
В настоящем исследовании показано, что воздействия, приводящие к увеличению сАМР в преадипоцитах, вызывают повышение [Са2+]i. Известно, что уровень сАМР в клетке определяется балансом между двумя процессами: процессом синтеза, опосредованным активацией аденилатциклазы, и процессом деградации, опосредованным активацией фосфодиэстеразы циклических нуклеотидов. Неспецифическим ингибитором фосфодиэстераз циклических нуклеотидов является соединение 3-изобутил-1-метилксантин (IBMX). Оптимальная концентрация IBMX для преадипоцитов бурого жира была выбрана в результате титрования вышеупомянутого ингибитора в широком диапазоне концентраций. Как следует из рисунка 5, 30 μМ является самой эффективной концентрацией. Таким образом, ингибирование фосфодиэстераз в присутствии IBMX вызывает увеличение уровня сАМР, что в свою очередь увеличивает кальциевый ответ на НА [1-6].
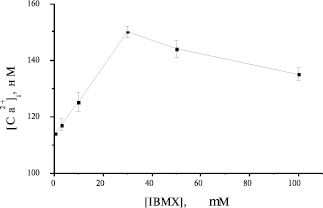
Рис. 5. Зависимость Са2+-ответа на НА от концентрации IBMX в преадипоцитах
Показано, что в адипоцитах бурого жира наиболее широко представлены фосфодиэстеразы 3- и 4-типов. Селективными ингибиторами фосфодиэстераз 3- и 4-типов являются, такие соединения как ОРС-3911 и Ro-20-1724, соответственно.
Ранее было показано, что ингибиторы фосфодиэстераз 3- и 4-типов синергично воздействовали на увеличение внутриклеточного уровня сАМР. Подобное явление наблюдалось также на культивируемых зрелых бурых адипоцитах. Совместное применение этих ингибиторов в данном исследовании на бурых преадипоцитах вызывало меньшее усиление Са2+-ответа на НА, чем вызывали эти ингибиторы по отдельности (рис. 6). На наш взгляд это полностью согласуется с колоколообразными кривыми дозозависимости эффекта BrcAMP (рис. 3) и IBMX (рис. 4) на величину Са2+-ответа в бурых преадипоцитах.
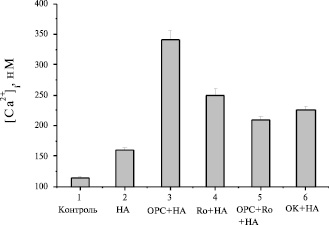
Рис. 6. Влияние ингибиторов фосфодиэстераз и фосфатазы на инициированный НА Ca2+-ответ клеток: 1 – контроль; 2 – 6µМ НА; 3 – 0,5 µМ OPC-3911+НА; 4 – 15 µМ Ro 20-1724+НА; 5 – OPC+Ro+НА; 6 – 30 µМ OK+НА
Известно, что активность Са2+-каналов зрелых клеток модулируется процессами фосфорилирования и дефосфорилирования, осуществляемыми протеинкиназой А и серин- треонин специфической фосфатазой. В данном исследовании установлено, что специфический ингибитор протеинкиназы А (Н-89) подавлял Са2+-ответы на изопротеренол дозозависимым образом (не показано) и форсколин. Максимальное подавление Са2+-ответа на форсколин ингибитором ПКА ([H-89]= 1 µM) составляло 55 % (рис. 7). Добавление ингибитора фосфатазы, окадаиковой кислоты, существенно увеличивало (рис. 6) Са2+-ответ на НА. Эти факты говорят в пользу того, что ПКА, фосфатаза и фосфодиэстераза участвуют в процессе формирования Са2+-ответа на норадреналин в бурых преадипоцитах.
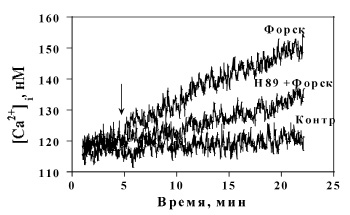
Рис. 7. Влияние ингибитора протеинкиназы А H-89 (1 μМ) на кинетику Ca2+-ответа, инициированного в бурых преадипоцитах форсколином (10 µM)
Таким образом, в данной работе показано, что Са2+-ответ в свежевыделенных бурых преадипоцитах инициируется главным образом через b-адренорецепторы и далее опосредуется через сАМР/протеинкиназа А-зависимый путь. Это является отличительной чертой бурых преадипоцитов. В зрелых адипоцитах, Са2+-ответы инициируются через α1-адренорецепторы.
Второй отличительной чертой Са2+-ответа в бурых преадипоцитах является колоколообразная зависимость его величины от концентрации агонистов адренорецепторов или сАМР; в зрелых клетках эти зависимости имели гиперболический или сигмоидальный вид. Согласно классическим представлениям колоколообразная дозозависимость в ответ на действие гормона или нейротрансмиттера указывает на его двойственное действие: он является активатором при низкой концентрации и ингибитором при высокой. Ранее были описаны подобные колоколообразные дозозависимости аккумуляции сАМР в зрелых бурых адипоцитах при аппликации норадреналина и изопротеренола. Ингибирующее действие высоких концентраций (1-10 μМ) НА и изопротеренола в этом случае было обусловлено активацией ими Са2+-сигнала. Возросший уровень [Ca2+]i приводил к активации СаМ/Са2+-зависимой фосфодиэстеразы и расщеплению сАМР. Колоколообразный характер кривых может быть связан с наличием отрицательной обратной связи, так например, показано на кардиомиоцитах, что при адренергической стимуляции L-типа Са2+-каналов увеличение [Ca2+]i приводит к подавлению аккумуляции сАМР, связанное в одном случае с Са2+- индуцированным ингибированием аденилатциклазы, в другом- с активацией СаМ-зависимой фосфодиэстеразы. Более того, с помощью моделирования показано, что наличие двух участков фосфорилирования Са2+-каналов в большей степени соответствует экспериментальным данным о двойственном эффекте изопротеренола на кардиомиоцитах [1-6].
Библиографическая ссылка
Мухаметдинова Э.С., Кужамбердиева С.Ж., Абжалелов Б.Б. МЕХАНИЗМ ФОРМИРОВАНИЯ КАЛЬЦИЕВОГО СИГНАЛА В ПРЕАДИПОЦИТАХ БУРОЙ ЖИРОВОЙ ТКАНИ // Международный журнал экспериментального образования. 2016. № 7. С. 59-65;URL: https://expeducation.ru/ru/article/view?id=10285 (дата обращения: 14.05.2026).