
Scientific journal
International Journal of Experimental Education
ISSN 2618–7159
ИФ РИНЦ = 0,827
EFFECT OF VOLATILE METABOLITES OF SAPROPHYTIC BACTERIA ISOLATED FROM DIFFERENT SOIL TYPES ON THE REPRODUCTION LISTERIA MONOCYTOGENES

Выживание патогенных бактерий, занимающих различные экологические ниши, может быть основано на симбиотическом взаимоотношении с различными биологическими объектами. Подобное взаимодействие патогенных микроорганизмов с представителями биоценоза различных естественных экосистем позволяет им сохраняться в сапрофитическом состоянии в межэпидемические периоды [4].
Одной из экологических ниш для патогенных бактерий является почва, имеющая свою биоту, составной частью которой является множество микроорганизмов с разнообразными биологическими свойствами. Жизнедеятельность микроорганизмов в почве осуществляется в основном на почвенных частицах, в определенных микрозонах которых представлены клетки, питательные вещества и микробные метаболиты.
Многие исследователи [1, 2] считают, что соединения, продуцируемые микроорганизмами, могут действовать как внутри- или межвидовые регуляторы микробных сообществ. Помимо веществ растворимых в воде, среди метаболитов, продуцируемых микроорганизмами, есть и летучие вещества [12]. При этом отмечено как стимулирующее, так и ингибирующее действие веществ микробного происхождения на размножение микроорганизмов [7, 12]. Изучение характера взаимоотношений между бактериями в микробоценозах имеет большое значение для выяснения роли отдельных компонентов в биологических сообществах.
Возможность того, что естественная микрофлора почвы может оказывать положительное влияние на сохранение патогенных микроорганизмов, представляет особый интерес для исследований в этом направлении. В отношении влияния почвенной микрофлоры на биологические свойства патогенов, входящих в состав микробного сообщества, имеются единичные работы [5, 7, 9].
Возбудитель листериоза, как представитель сапрозоонозной инфекции, длительно сохраняется и размножается в почвенных экосистемах [3, 11].
В настоящее время установлено, что на жизнеобитание этих бактерий в почвах оказывают влияние как абиотические факторы среды [6, 10], так и биотические [7, 8]. Влияние метаболитов сапрофитных почвенных бактерий, ассоциированных с листериями в почвенных сообществах, на размножение Listeria monocytogenes практически не изучено.
Цель исследования. Цель настоящей работы – изучить влияние летучих метаболитов сапрофитных бактерий, выделенных из микробоценозов разных типов почв, на размножение Listeria monocytogenes.
Материалы и методы исследования
При проведении экспериментальных исследований в качестве объектов исследования были использованы штаммы сапрофитных бактерий, выделенные из разных типов почв: бурая лесная типичная, бурая лесная оглеенная, дерново-подзолистая, буро-подзолистая. Почвенные образцы отбирали из верхнего (0-10 см) горизонта.
Для каждого типа почвы были определены указанные показатели (табл. 1).
Таблица 1
Физико-химические свойства почвенных образцов
|
Показатели тип почвы |
рН |
Гумин, % |
С, % |
|
бурая лесная типичная |
6,5 |
17,39 |
2,0 |
|
дерново-подзолистая |
5,4 |
30,5 |
2,5 |
|
буро-подзолистая |
7,3 |
22,14 |
2,6 |
|
бурая лесная оглеенная |
5,5 |
10,4 |
1,1 |
Выбор представленных биотопов объясняется тем, что наибольшее количество выделений L. monocytogenes отмечается в почвах лесной зоны (Гершун, 1988; Сомов, Литвин, 1988). Данные биотопы представляют высокий риск в распространении листериозной инфекции.
В качестве тест-микроорганизмов для исследования были использованы штаммы Listeria monocytogenes, типичные по своим культуральным, серологическим и биохимическим свойствам, взятые из музея «НИИЭМ имени Г.П. Сомова» СО РАМН.
Сапрофитные бактерии выделяли и выращивали для экспериментов на пептонном агаре (ПА) (1 % пептона, 0,5 % NaCI на дистиллированной воде с 2 %-м агаром, рН=7,4) при температуре 22 °С, 20-24 ч. Листерии культивировали на питательном агаре с дрожжевым экстрактом и 0,1 % глюкозой при температуре 37 °С, 20-24 ч.
Морфотипы сапрофитных бактерий выделяли по морфологическим и культуральным признакам колоний. Микроскопию, окрашенных по Граму мазков, проводили на микроскопе Axio Scope A1 («Carl Zeiss», Германия).
Для определения биологической активности летучих метаболитов использовали скрининговый экспресс-метод Л.С. Тирранен (1980) в нашей модификации. Культуру сапрофитных бактерий высевали газоном на чашки Петри с питательным агаром. На чашки Петри с казеиново-дрожжевым агаром производили посев уколом культуры листерий. Чашки с посевами сапрофитов и листерий соответственно состыковывали. Посевы выдерживали в течение семи суток при комнатной температуре.
Для оценки размножения L. monocytogenes замеряли зоны роста в месте укола. Воздействие культуры сапрофитов на тест-культуру L. monocytogenes оценивали как положительное (стимулирующее) или отрицательное (ингибирующее), когда размер колоний тест-культур в опыте был соответственно увеличен или снижен на 20 % и более по сравнению с контролем. Если размер колоний тест-культуры L. monocytogenes в опыте отличался от контрольной не более чем на ±20 %, действие испытуемой культуры оценивали как нулевое. Долю случаев положительного влияния летучих метаболитов на листерии выражали в процентах (%). Критерием отбора активных штаммов, проявивших максимальную стимуляцию размножения листерий, служил диаметр зон роста листерий больше 10 мм (d ≥ 10).
Все опыты проводились в трехкратной повторности. Статистическую обработку результатов проводили с помощью программы Microsoft Office Excel 2007.
Результаты исследования и их обсуждение
Всего было выделено 135 морфотипов сапрофитных бактерий, различных по своим культуральным и морфологическим признакам (табл. 2).
Таблица 2
Биологическая активность и численность морфотипов сапрофитных бактерий, выделенных из разных типов почв
|
Источник выделения |
Общее количество штаммов |
Доля случаев положительного влияния на рост листерий, % |
Количество активных штаммов |
Количество штаммов, активных в отношении L. monocytogenes, серотип 4b |
|
бурая лесная типичная |
53 |
76,5 |
7 |
1 |
|
дерново-подзолистая |
23 |
48,3 |
6 |
2 |
|
буро-подзолистая |
35 |
55,7 |
10 |
3 |
|
бурая лесная оглеенная |
24 |
32,5 |
5 |
3 |
Высокие показатели численности морфотипов (табл. 2), выделенных из бурой лесной типичной и буро-подзолистой объясняются тем, что для данных типов почв характерен богатый растительный опад, преимущественно широколиственных пород, слабокислая реакция среды, высокая насыщенность основаниями и достаточное количество легкоокисляемой органики, что способствуют интенсивному развитию в них микроорганизмов и многообразию видов.
Меньшая численность морфотипов (в 2 раза меньше по сравнению с бурой лесной типичной), выделенных из дерново-подзолистой почвы объясняется малой мощностью дернового горизонта и кислой реакцией (рН=5,4), которая неблагоприятна для многих микроорганизмов.
Численность морфотипов, выделенных из бурой лесной оглеенной почвы, также низка, по сравнению с бурой лесной типичной. Это объясняется типичным для бурой лесной оглеенной почвы рядом характеристик. Процессы оглеения, происходящие в бурой лесной оглеенной почве, связаны с избыточным переувлажнением, развитием восстановительных процессов в анаэробных условиях, подкислением реакции почвы, что неблагоприятно отражается на качественной и количественной структуре микрофлоры эти почв.
Если рассматривать показатели биологической активности выделенных штаммов на тест-культуры, то видно, что сапрофитная микрофлора бурых лесных типичных почв оказывает наибольшее стимулирующее влияние на листерии, по сравнению с сапрофитными бактериями, выделенными из остальных типов почв (буро-подзолистая, дерново-подзолистая, бурая лесная оглеенная) (рисунок 1, а). При этом наибольшее количество активных штаммов выделено из буро-подзолистой почвы (табл. 2).
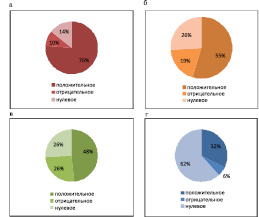
Оценка влияния летучих метаболитов сапрофитных бактерий, выделенных из разных типов почв, на размножение листерий, ( % случаев): а – бурая лесная типичная, б – буро-подзолистая, в – дерново-подзолистая, г – бурая лесная оглеенная
Штаммы, выделенные из бурой лесной оглеенной почвы оказывали стимулирующее влияние в 32 % случаев (рис. 1, г). На диаграмме показано, что большинство случаев характеризовались как нулевые, они могли быть слабо отрицательными или слабо положительными. Но, несмотря на меньшее число морфотипов и количество активных штаммов из бурой лесной оглееенной почвы, они в большей степени проявили биологическую активность в отношении эпидемически значимого штамма L. monocytogenes, серотип 4b (табл. 2).
Результаты сравнительной характеристики показывают зависимость количества морфотипов бактерий и их биологической активности от источника выделения (типы почв). Выявлено, что показатели биологической активности выше у штаммов, выделенных из почв с неблагоприятными для микроорганизмов условиями.
Выводы
Таким образом, отмечено стимулирующее, ингибирующее и нулевое действие летучих метаболитов сапрофитных почвенных бактерий. Количественная оценка влияния летучих метаболитов на размножение листерий, показала, что в большинстве случаев преобладает положительное действие. Так же выявлено, что численность морфотипов бактерий, а также их биологическая активность в отношении листерий зависела от условий типа почв из которых они выделены. Большую степень биологической активности проявили штаммы из почв с неблагоприятными для микроорганизмов условиями.
Библиографическая ссылка
Ли Н.Г., Бузолева Л.С., Сидоренко М.Л. ВЛИЯНИЕ ЛЕТУЧИХ МЕТАБОЛИТОВ САПРОФИТНОЙ МИКРОФЛОРЫ РАЗНЫХ ТИПОВ ПОЧВ НА РАЗМНОЖЕНИЕ LISTERIA MONOCYTOGENES // Международный журнал экспериментального образования. 2013. № 10-2. С. 288-291;URL: https://expeducation.ru/en/article/view?id=4231 (дата обращения: 16.07.2026).