
Scientific journal
International Journal of Experimental Education
ISSN 2618–7159
ИФ РИНЦ = 0,827
PATTERNS OF SPATIO-TEMPORAL DYNAMICS OF MAMMALS’ POPULATIONS IN THE EASTERN PART OF MIDDLE URALS (SVERDLOVSKAYA OBLAST’)

Анализ биологического разнообразия, как одного из основных факторов, определяющих устойчивость природных сообществ к внешним воздействиям (например, природным катастрофам) требует выделения комплексов видов, со схожими функциональными характеристиками. Существует несколько подходов к решению данной проблемы. Один из них состоит в том, что виды объединяются в функциональные группы на основе имеющихся a priori представлений об их таксономическом положении, трофических связях, размерных характеристиках – на этом построены исследования, соответственно, таксономического, функционального и морфологического разнообразия. Альтернативой такому подходу может служить объединение видов в группы по признаку, определенному a posterior, по их реакции на воздействие комплекса факторов окружающей среды. Такой подход требует анализа синхронности в изменениях численности различных видов. В более общем виде, данная проблема сводится к вопросу о пространственной корреляции популяционной динамики вида или видов [9], то есть о связи синхронности колебаний численности на разных участках, расположенных на различных расстояниях друг от друга. Авторы, главным образом, используют синхронные колебания обилия видов для проверки гипотез о влиянии на них какого-либо фактора, например, погодных условий или общего хищника [8]. Такой подход, по сути, является классическим для экологии животных. В то же время, большой интерес представляет и обратная постановка вопроса: можно ли по синхронным колебаниям численности разных видов (или одного вида на разных территориях) оценить какие именно факторы играют ведущую роль в формировании популяционной динамики?
В данном исследовании мы анализируем пространственно-временные закономерности населения ряда видов млекопитающих восточной части Среднего Урала (Свердловская область) с целью выявления групп экологически сходных видов и факторов, определяющих их обилие.
Уральский регион характеризуется высоким разнообразием ландшафтов, что обусловливает особенности видового состава животных в разных его частях. Структура населения млекопитающих Свердловской области рассматривалась ранее в работах В.Н. Большакова с соавторами [2, 3]. В первой из упомянутых работ описаны «териографические провинции», которые характеризуются «ключевыми» видами млекопитающих, маркирующими различные ландшафты. Эта классификация была основана на качественных оценках распространения и ключевых экологических характеристиках зверей. Во второй работе был использован количественный подход – на основании многолетних данных о численности лося в административных районах Свердловской области за 20 лет были описаны территории со сходной динамикой численности животных. Границы этих областей были весьма сходны с границами лесорастительных областей по классификации Б.П. Колесникова [6].
Объектом нашего исследования является группа видов млекопитающих, которые отнесены к охотничьим ресурсам и учитываются с помощью метода зимнего маршрутного учета [7]. В ходе работы проверяли, в частности, гипотезы о существовании внутри региона областей, различающихся по обилию исследуемых видов, а также о существовании пространственно-временной когерентности в динамике их населения в различных природных зонах (в пределах Свердловской области).
Материалы и методы исследования
В качестве источника информации о численности охотничьих зверей были использованы данные зимних маршрутных учетов, проведенных в период с 1989 по 2008 гг. государственными службами, отвечающими за охрану и планирование использования охотничьих ресурсов. Из всего спектра видов, попадающих в данный вид учетов, были выбраны те, для которых оценки обилия, полученные с помощью ЗМУ, отличаются относительно высокой достоверностью: белка (Sciurus vulgaris), горностай (Mustela erminea), заяц-беляк (Lepus timidus), кабан (Sus scrofa), сибирская косуля (Capreolus pygargus), лесная куница (Martes martes), лисица (Vulpes vulpes), лось (Alces alces). База данных в исходном варианте представляла собой сведения о численности животных в 45 административных районах области за каждый год для периода с 1980 по 2008 год, то есть за 20 лет. Эти сведения, на основании информации о площади охотничьих угодий в каждом из районов области, были трансформированы в показатели плотности населения, выраженной как число особей на 1000 га угодий. Далее, показатели плотности были, с целью параметризации, трансформированы по формуле Y=(ln x +1). Итогом расчетов стала полученная для каждого вида матрица данных размером 45 (районов) х 20 (лет). Эти матрицы были проанализированы с помощью методов кластерного анализа (мера дистанции – квадрат Эвклидова расстояния между объектами, связывание по методу Уорда). В результате были выделены группы районов со сходной динамикой плотности населения животных каждого вида.
При выделении групп районов мы использовали максимально возможную степень генерализации – выделяли кластеры последнего (максимальная дистанция объединения) или предпоследнего (дистанция объединения как минимум в два раза выше, чем у нижележащих кластеров) ранга. Далее, для каждой из выделенных групп районов мы рассчитали среднюю плотность населения животных за каждый год наблюдений. Полученные временные тренды динамики обилия были сопоставлены между собой – проверяли гипотезы о наличии значимых различий в показателях обилия между различными группами районов, а также о синхронности колебаний плотности населения животных в них. Для проверки гипотез использовали стандартные статистические индексы: t-критерий Стьюдента, критерий Манна-Уитни, коэффициенты корреляции Пирсона и Спирмана.
Результаты исследования и их обсуждение
Анализ пространственного размещения районов, сходных по динамике плотности населения животных каждого вида позволил выявить следующие закономерности их численности и пространственного размещения (см. здесь и далее табл. 1 и рис. 1):
Таблица 1
Плотность населения исследуемых видов в выделенных «кластерах» районов
|
Группа 1 |
|||
|
Вид |
«Северный кластер» |
«Южный кластер» |
|
|
Лисица |
0,18 |
0,53 |
|
|
Горностай |
1,0 |
0,25 |
|
|
Колонок |
0,67 |
0,28 |
|
|
Группа 2 |
|||
|
«Западный кластер» |
«Восточный кластер» |
||
|
Заяц-беляк |
9,6 |
4,68 |
|
|
Лось |
1,9 |
0,91 |
|
|
Группа 3 |
|||
|
«Юго-восток» |
«Центр» |
«Северо-восток» |
|
|
Косуля |
4.63 |
0.75 |
0.065 |
|
Кабан |
0.57 |
0.27 |
0.077 |
|
Белка |
0,94 |
5,55 |
15,45 |
1. Население таких видов, как колонок и горностай в Свердловской области разбивается на два кластера – северный и южный. Вектор уменьшения плотности направлен в сторону низких широт. Как для горностая, так и для колонка анализ выявил районы относительно низкой плотности в северной части области. Аналогично, на юге имеются отдельные районы, для которых показатели плотности населения вида выше, чем на соседних территориях. Размещение таких «островных» участков внутри северного и южного кластеров не имеет четкой географической закономерности, однако примечательно, что у обоих видов они располагаются в одних и тех же, или близко расположенных районах. Также к этой группе может быть отнесена лисица, однако у нее, численность, напротив, оказывается выше в южных районах.
2. Для таких видов как лось и заяц-беляк территория области может быть разбита также на два кластера, но здесь дифференциация населения идет, главным образом, в меридиональном направлении. Численность указанных видов в западной, горной, части области в целом выше, чем в восточной, равнинной, ее части. В то же время, для зайца-беляка в кластер с высокой численностью включаются также и некоторые южные районы области. Обратим внимание, что если на большей части области численность лося и зайца варьирует сходным образом, то в юго-восточных районах она носит разнонаправленный характер: отдельные районы юго-востока, которые имеют высокую численность лося, характеризуются низкой численностью зайца беляка, и, наоборот, в районах где плотность населения беляка относительно высока, лось немногочисленен. Исключение составляет Тугулымский район, располагающийся на крайнем юго-востоке области – здесь оба вида характеризуются высокой плотностью населения.
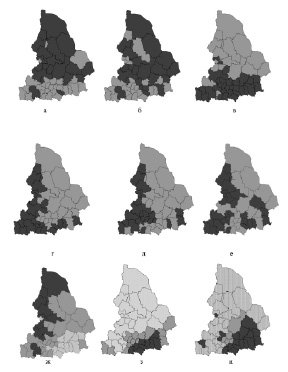
Результаты кластерного анализа динамики численности охотничьих животных в административных районах Свердловекой области. Более темным цветом показаны районы с более высокой плотностью населения. Виды: а – горностай; б – колонок; в – лисица; г – заяц-беляк; д – лось; е – куница; ж – белка; з – сибирская косуля; и – кабан
3. Для кабана, косули, белки территория области по обилию животных может быть разбита на три кластера, один из которых располагается на юго-востоке региона, другой охватывает центральную часть области, третий включает районы северо-западной его части. У первых двух видов численность снижается в направлении от юго-востока к северо-западу, для белки отмечена противоположная тенденция.
Лесная куница не демонстрирует четких географических закономерностей вариации численности.
Когерентность динамики обилия животных между выявленными кластерами районов была проанализирована для каждого из исследованных видов. Для тех, у которых было описано два кластера (горностай, колонок, заяц, лось), показано высокое сходство кривых многолетней динамики плотности населения в обоих кластерах, вне зависимости от того имела ли место дифференциация населения в направлении «север-юг» или «запад-восток» (табл.2). Более того, корреляция динамики численности наблюдалась не только внутри кластеров, но и между ними, например, для динамики населения колонка в северном кластере наблюдает статистически значимая корреляция не только с численностью горностая не только на севере, но и на юге региона, а также высока корреляция между северным и южным кластерами. Другими словами, для указанных видов не удалось выявить значимых различий в динамике населения животных в разных пространственных кластерах. Таким образом, можно предполагать, что выявленная кластерная структура населения была обусловлена только различиями в численности животных на этих территориях, но не характером популяционной динамики.
Таблица 2
Матрица коэффициентов корреляции Спирмана R динамики обилия видов в выделенных географических кластерах (группы 1 и 2).
|
Вид |
Кластер |
Группа 1 |
|||
|
Горностай |
Колонок |
||||
|
север |
юг |
север |
юг |
||
|
горностай |
северный |
– |
0.59 |
0.726 |
0.279 |
|
южный |
– |
0.887 |
0.843 |
||
|
колонок |
северный |
– |
0.767 |
||
|
южный |
– |
||||
|
группа 2 |
|||||
|
заяц-беляк |
лось |
||||
|
запад |
восток |
запад |
восток |
||
|
заяц-беляк |
западный |
– |
0.792 |
0.445* |
0.65 |
|
Восточный |
– |
0.478* |
0.592 |
||
|
лось |
западный |
– |
0.634 |
||
|
восточный |
– |
||||
Для видов, у которых было выделено три кластера (белка, кабан, косуля) матрица коэффициентов корреляции всего массива данных также не показала четких различий в характере динамики численности между географическими кластерами (данные не представлены). В то же время, при анализе связи показателей обилия между видами внутри отдельных кластеров прослеживаются определенные закономерности. Так, в юго-западной части области наблюдалась положительная корреляция обилия всех трех видов. В центральных районах связь наблюдалась только для кабана и косули, а в северо-западных районах корреляция между численностью рассматриваемых видов была статистически незначима (табл. 3).
Таблица 3
Значения коэффициентов корреляции Спирмана динамики обилия видов группы 3
|
Вид |
Географический кластер |
Вид |
|
|
косуля |
белка |
||
|
кабан |
Юго-восточный |
0.61** |
0.54* |
|
Центральный |
0.55* |
-0.39* |
|
|
Северо-западный |
0.26 |
-0.15 |
|
|
косуля |
Юго-восточныйв |
0.48* |
|
|
Центральный |
-0.17 |
||
|
Северо-западный |
0.1 |
||
Обсуждение. Рассмотренные виды млекопитающих могут быть объединены в группы по характеру пространственной динамики плотности их населения. Это говорит, во-первых, о наличии природных факторов, определяющих крупномасшатбную динамку населения животных, а во-вторых, о том, что данные факторы оказывают влияние не на одиночные виды, а на комплексы видов. Количественный анализ влияния данных факторов на животных выходит за рамки данной работы. Мы сформулируем предположения, относительно того, какие характеристики местообитаний могут определять наблюдаемую картину распределения зверей.
Наличие широтной динамики населения предполагает влияние климатических показателей. При рассмотрении вариации обилия в меридиональном направлении обращает на себя внимание различия в характере рельефа: западная часть характеризуется горным, тогда как восточная – равнинным рельефом. Однако, известно, что такие факторы как климат и рельеф редко оказывают прямое действие на животных [1] . Наблюдаемые изменения обилия могут быть обусловлены, скорее, действием комплекса параметров среды обитания, из которых важнейшими можно полагать структуру ландшафтов, включая особенности растительного покрова, и характер антропогенного воздействия, как на самих животных, так и на среду их обитания. Поэтому, выделенные нами группы видов могут быть интерпретированы следующим образом: виды, реагирующие на «широтные» факторы (группа 1), виды, реагирующие на «долготные факторы» (группа 2) и виды, реагирующие на сочетание «широтных» и долготных» факторов.
В то же время, важно отметить, что сами по себе «широтные» и «долготные» факторы определяют лишь различия в плотности населения животных, и для групп 1 и 2 направления изменений численности одинаковы в разных природных зонах. Это позволяет предполагать, что ключевое влияние оказывают, скорее, не абиотические факторы, такие как рельеф и климат, а изменения состояния биоты, например, характер кормовой базы, воздействие хищников или других факторов смертности.
Группа 1 образована хищными млекопитающими. Для них в качестве регулятора численности могут выступать изменения обилия грызунов, которые происходят синхронно на больших территориях [5]. В состав группы 2 входят растительноядные животные – лось и заяц-беляк. Вопрос о причинах синхронных колебаний их обилия остается открытым. Некоторые авторы рассматривают в качестве возможной причины многолетних колебаний численности растительноядных животных, изменения продуктивности растительных сообществ под влиянием динамики солнечной активности [10]. Другая гипотеза состоит в превалирующей роли социально-экономических факторов, в частности, уровня незаконной добычи [4] или масштабов вырубки лесов. В случае Свердловской области, такие причины могут оказывать влияние главным образом на динамику численности лося [3], для зайца-беляка этот вопрос требует дополнительных исследований. Группа 3 образована также растительноядными видами. Особенностями видов этой группы является то, они не являются эвритопами, и в пределах региона для каждого из них отчетливо выделяются зоны с оптимальными и пессимальными условиями обитания. В частности, для кабана и косули зона экологического оптимума располагается на юго-востоке области, а самые неблагоприятные условия имеются в северо-западной ее части, для белки наблюдается прямо противоположная ситуация [2, 3]. Зоны экологического оптимума и пессимума обусловлены здесь комплексным воздействием трофических, защитных и погодных факторов, что, скорее всего, и определяет характер варьирования обилия как в широтном, так и в долготном аспекте.
Анализ корреляции динамики обилия видов внутри и между географическими кластерами показал, что ключевым параметром, определяющим существование таких кластеров, являются только различия в уровне обилия животных, но не форма кривых изменения численности. Таким образом, предполагаемые факторы, определяющие обилие зверей, действуют более или менее синхронно, по крайней мере, в масштабе области.
В целом, наши результаты позволяют предполагать ведущую роль трофического фактора в формировании пространственно-временной структуры населения охотничьих зверей района исследований.
Работа выполнена при поддержке проекта 12-С-4-1012 УрО РАН «Пространственно-временная когерентность экологических процессов на территории Евразии».
Библиографическая ссылка
Марков Н.И., Жигальский О.А. ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННО-ВРЕМЕННОЙ ДИНАМИКИ НАСЕЛЕНИЯ МЛЕКОПИТАЮЩИХ ВОСТОЧНОЙ ЧАСТИ СРЕДНЕГО УРАЛА (СВЕРДЛОВСКАЯ ОБЛАСТЬ) // Международный журнал экспериментального образования. 2013. № 10-2. С. 292-297;URL: https://expeducation.ru/en/article/view?id=4232 (дата обращения: 16.07.2026).